|
мать и отец обладают нормальным зрением, половина сыновей может оказаться
пораженными цветовой слепотой. В основном наличие цветовой слепоты чаще
встречается у мужчин. Э.Вильсон объяснил наследование этого признака,
предположив, что он локализовал в Х-хромосоме и что у человека гетерогаметным
(XY) является мужской пол. Становится вполне понятным, что в браке гомозиготной
нормальной женщины (Ха Ха) с мужчиной дальтоником (Х
аy) все дети рождаются нормальными. Однако при этом, все дочери становятся
скрытыми носителями дальтонизма, что может проявиться в последующих поколениях.
Другим примером наследования сцепленного с полом, может послужить
рецессивныйполулетальный ген, вызывающий несвертываемость крови на воздухе -
гемофилию. Это заболевание появляется почти исключительно только у мальчиков.
При гемофилии нарушается образование фактора VIII, ускоряющего свертывание
крови. ген, детерминирующий синтех фактора VIII, находится в участке Х-
хромосомы, недоминантным нормальным и рецессивным мутантным. Возможны
следующие генотипы и фенотипы:
Генотипы | Фенотипы | Хн Хн | Нормальная женщина | Хн Хn | Нормальная женщина (носитель) | Хнy | Нормальный мужчина | Хny | Мужчина гемофилик |
В гомозиготном состоянии у женщин ген гемофилии летален.
Особей женского пола, гетерозиготных по любому из сцепленных с полом
признаков, называют носителями соответствующего рецессивного гена. Они
фенотипически нормальны, но половина их гамет несет рецессивный ген. Несмотря
на наличие у отца нормального гена, сыновья матерей-носителей с вероятностью
50% будут страдать гемофилией.
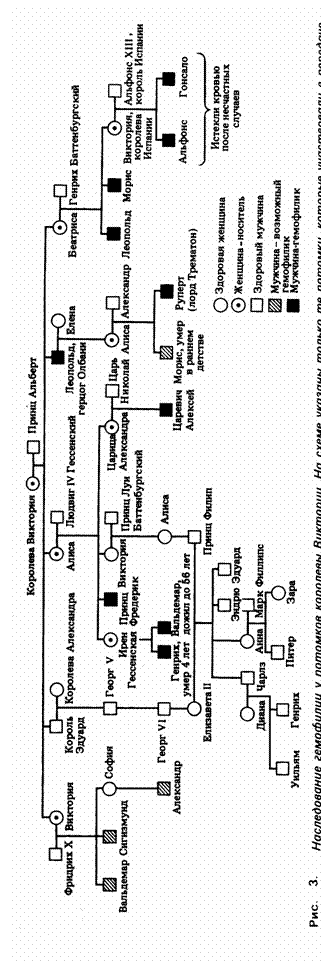
Один из наиболее хорошо документированных примеров наследования гемофилии мы
находим в родословной потомков английской королевы Виктории. Предполагают,
что ген гемофилии возник в результате мутации у самой королевы Виктории или
у одного из ее родителей. Среди унаследовавших это врожденное заболевание -
цесаревич Алексей, сын последнего русского царя Николая II. Мать цесаревича,
царица Александра Федоровна (Алиса, рис.2), получила от своей бабушки
королевы Виктории ген гемофилии и передала его в четвертом поколении бывшему
наследнику царского престола. На рис.2 показано, как этот ген передавался ее
потомкам.

Один из сцепленных с полом рецессивных генов вызывает особый тип мышечной
дистрофии (тип Дюмена). Эта дистрофия проявляется в раннем детстве и
постепенно ведет к инвалидности и смерти ранее 20-летнего возраста. Потому
мужчины с дистрофией Дюмена не имеют потомства, а женщины гетерозиготные по
гену этого заболевания, вполне нормальны.
Среди доминантных признаков, связанных с Х-хромосомой, можно указать на ген,
который вызывает недостаточность органического фосфора в крови. В результате,
при наличии этого гена, часто развивается рахит, устойчивый к лечению
обычными дозами витамина А. В этом случае картина сцепленного с полом
наследования заметно отличается от того хода передачи по поколениям, который
был описан для рецессивных болезней. В браках девяти больных женщин со
здоровыми мужчинами среди детей была половина больных девочек и половина
мальчиков. Здесь, в соответствии с характером наследование доминантного гена,
в Х-хромосомах произошло расщепление в отношении 1:1:1:1.
Другим примером доминантного гена, локализованного в Х-хромосоме человека,
может послужить ген, вызывающий дефект зубов, приводящий к потемнению эмали
зубов.
Так как гетерогаметный пол гемизиготен по сцепленным с полом генам, то эти
гены всегда проявляются в их фенотипе, даже если они рецессивны. Большинство
генов, имеющихся в Х-хромосоме, в Y-хромосоме отсутствует, однако
определенную генетическую информацию она все-таки несет. Различают два типа
такой информации: во-первых, содержащуюся в генах, присутствующих только в Y-
хромосоме, и, во-вторых, в генах, присутствующих как в Y-, так и в Х-
хромосоме (гемфрагический диатез).
Y-хромосома передается от отца всем его сыновьям, и только им. Следовательно,
для генов, содержащихся только в Y-хромосоме, характерно голандрическое
наследование, т.е они передаются от отца к сыну и проявляются у мужского
пола.
У человека в Y-хромосоме содержатся по крайней мере три гена, один из
которых необходим для дифференциации семенников, второй требуется для
проявления антигена гистосовместимости, а третий оказывает влияние на размер
зубов. Y-хромосома имеет немного признаков, среди которых есть
патологические. Патологические признаки наследуются по параллельной схеме
наследования (100%-ое проявление по мужской линии). К ним относят:
1) облысение;
2) гипертрихоз (оволосенение козелка ушной раковины в зрелом возрасте);
3) наличие перепонок на нижних конечностях;
4) ихтиоз (чешуйчатость и пятнистое утолщение кожи).
2.3. Наследование признаков, контролируемых полом.
Имеется ряд признак, контролируемых генами, расположенными в аутосомах,
однако для проявления этих признаков необходима определенная среда,
создаваемая генами, находящимися в половых хромосомах (например, гены,
определяющие мужские признаки, находятся в аутосомах, и их фенотипические
эффекты маскируются наличием пары Х-хромосом, в присутствии одной Х-хромосомы
мужские признаки проявляются. Такие признаки называются обусловленными или
контролируемыми полом. Появление лысины - аутосомно-доминантный признак, но
проявляется практически только у мужчин при наследовании, контролируемом
полом, у женщин подавляются гены, детерминирующие рост бороды.
3. Сцепленное наследование признаков.
Наряду с признаками, наследуемыми независимо, обнаружены признаки,
наследуемые совместно (сцепленно). Экспериментальное наследование этого
явления, проведенное Т.Г. Морганом и его группой (1910-1916), подтвердило
хромосомную локализацию генов и легло в основу хромосомной теории
наследственности.
3.1. Хромосомная теория наследственности.
В работах на плодовой мушке Drosophila melanogaster было установлено, что гены
по признаку совместной их передачи потомкам подразделяются на 4 группы. Число
таких групп сцепления равно количеству хромосом в гаплоидном наборе. Можно
заключить, что развитие признаков, которые наследуются сцепленно,
контролируется генами одной хромосомы. Этот вывод обосновывается также данными
следующих наблюдений. Скрещивание серой мухи (В) с нормальными крыльями (V) и
черной мухи (в) с зачаточными крыльями (v) дает в 1-ом поколении серых гибридов
с нормальными крыльями 
. При скрещивании самца-гибрида 1-го поколения с черной самкой с зачаточными
крыльями  рождаются рождаются
особи 2 видов, аналогичных исходным родительским формам, причем в равном
количестве.
Полученные в проведенных скрещиваниях данные нельзя объяснить независимым
наследованием признаков. Рассматриваемые совместно результаты обоих
скрещиваний убеждают в том, что развитие альтернативных признаков
контролируется различными генами, и сцепленное наследование этих признаков
объясняется локализацией генов в одной хромосоме.
Основные положения хромосомной теории наследственности, сформулированной Т.Г.
Морганом, заключаются в следующем.
1. Гены располагаются в хромосомах; различные хромосомы содержат неодинаковое
число генов каждой из негомологичных хромосом уникален.
2. Аллельные гены занимают определенные и идентичные локусы гомологичных
хромосом.
3. В хромосоме гены располагаются в определенной последовательности по ее
длине в линейном порядке.
4. Гены одной хромосомы образуют группу сцепления, благодаря чему имеет место
сцепленное наследование некоторых признаков; сила сцепления находится в
обратной зависимости от расстояния между генами.
5. каждый биологический вид характеризуется специфичным набором хромосом
кариотипом.
3.2. Механизм сцепления.
Гены, локализованные в одной хромосоме, называют группой сцепления. Число
групп сцепления соответствует гаплоидному набору хромосом.
Если две сцепленные пары генов находятся в одной гомологичной паре хромосом, то
генотип запишется  . .
 Проведем скрещивание двух Проведем скрещивание двух
организмов различающихся по двум парам признаков, например
  Р Р  х х 
F1 
 Скрещивая гибриды 1-го поколения, получим Скрещивая гибриды 1-го поколения, получим
   х х 
F1
1 :2 :2 :1 :1 . .
Гены, находящиеся в одной паре гомологичных хромосом, наследуются вместе и не
расходятся в потомстве, так как при гаметогенезе они обязательно попадают в
одну гамету. Совместное наследование генов, ограничивающее свободное их
комбинирование называют сцеплением генов. Для наследования сцепленных генов,
находящихся в половых хромосомах, имеет значение направление скрещивания.
Нужно иметь ввиду, что кроме истинного сцепления, могут встречаться явления,
внешне сходные со сцеплением, нот отличные от него по природе: это так
называемое ложное, межхромосомное сцепление, возникающее из-за нарушения
свободного комбинирования негомологичных хромосом в мейозе. Такие случаи
наблюдались в скрещиваниях линий лабораторных мышей и дрожжей.
Страницы: 1, 2, 3
| 
